
中国滨海湿地的蓝色碳汇功能及碳中和对策
|
|
中国网/中国发展门户网讯 滨海湿地是由沿海盐沼湿地和红树林组成的湿地生态系统。由于受到海水周期性潮汐淹没的影响,滨海湿地的碳汇功能强大,是降低大气二氧化碳(CO2)浓度、减缓全球气候变化的重要途径。这些滨海湿地与海草床等生态系统所固存的碳被称为海岸带“蓝碳”(Blue Carbon)。“蓝碳”是与陆地植被的“绿碳”相对而言的。海洋每年从大气中净吸收(进出通量之差)大约 2.3Pg C·a-1,而陆地生态系统大约净吸收了 2.6Pg C·a-1 。传统上,海洋蓝碳被认为主要通过物理溶解度泵(大气 CO2 溶解到海水里)、生物泵(植物通过光合作用吸收和转化 CO2 并沉积到海底),以及海洋碳酸盐泵(贝类、珊瑚礁等海洋生物对碳的吸收、转化和释放)在不同时间尺度实现储碳。根据联合国的评估,全球海洋活体生物所固持的碳有一半位于海岸带的蓝碳生态系统。滨海湿地作为一类重要的海岸带蓝碳生态系统,具有巨大的碳吸收能力,属于“基于自然的解决方案”的实践范畴,是重要的基于海洋的气候变化治理手段之一;在减缓温室气体排放的同时,滨海湿地可以给沿海国家乃至全球带来经济和社会效益。研究表明,滨海湿地每平方公里的年碳埋藏量预计可达 0.22 Gg C,相当于燃烧 3.36×105 L 汽油燃烧所排放的 CO2。因此,有效地评估滨海湿地的碳汇能力、固碳潜力和生态系统服务功能,是制定减排增汇措施的重要手段,也是各国政府制定应对气候变化行动计划的理论依据,更是我国实现碳中和目标的重要基础。
滨海湿地的蓝碳固碳功能及机制
滨海湿地生态系统相比于陆地生态系统的优势在于极大的固碳速率,以及长期持续的固碳能力。陆地生态系统随植物的不断生长和土壤有机质的累积,其植物和土壤呼吸释放的碳会持续增加。因而,其固碳能力在几十年到百年尺度上会达到饱和。达到饱和点后,植物通过光合作用吸收的碳与系统内植物、微生物和动物呼吸释放的碳会达到平衡,从而导致系统净固碳能力趋于零。滨海湿地中植物的凋落物会沉积到土壤中,但是与陆地生态系统不同的是:海水潮汐往复能够极大减缓这些沉积有机质的分解;随着海平面的上升,滨海湿地中沉积物不断增加并被埋藏到更深的土层,客观上不利于有机质的降解,因而这些沉积物中的碳能够在百年到上万年尺度上处于稳定状态而不会释放回大气中,从而实现稳定持续的储碳。此外,与淡水湿地相比,由于海水中大量硫酸根离子的存在,能够有效抑制滨海湿地中的甲烷(CH4)排放。滨海湿地的这些特性相对于陆地生态系统固碳具有明显的优势,其单位面积的碳埋藏速率是陆地森林系统的几十到上千倍。故而长期来看,滨海湿地生态系统比陆地生态系统具备更强的固碳能力和生态系统服务功能,是应对人类目前所面临的气候变化问题的重要资源。
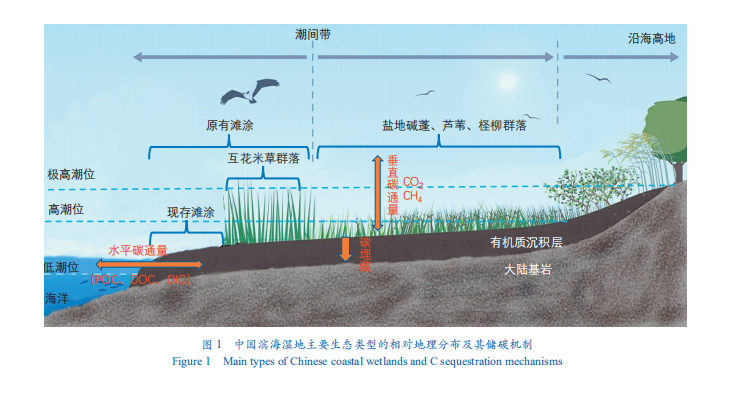
滨海湿地固碳功能主要体现在其垂直方向的沉积物的碳埋藏速率和水平方向通过潮汐作用与海水中的无机碳(DIC)、溶解有机碳(DOC)和颗粒有机碳(POC)的交换(图 1)。全球尺度上,盐沼湿地面积约为 6.23×104km2,红树林面积为 14×104km2 。根据滨海盐沼湿地和红树林的全球分布,结合过去在海岸带地区的有机质沉积速率的研究,Wang 等初步估算了全球尺度上盐沼湿地和红树林的碳埋藏速率约为 53.65Tg C·a-1,换算成 CO2 当量为 196.71Tg·a-1。这一数据相当于人类活动每年排放量的 0.6%。如果从单位面积碳埋藏速率估算,滨海湿地蓝碳系统的碳埋藏速率是陆地生态系统固碳速率的 15 倍、海洋生态系统固碳速率的 50 倍左右。因此,滨海湿地蓝碳系统的碳埋藏速率很高。此外,此数值仅为垂直方向碳埋藏速率,其水平方向通过潮汐作用与海洋的交换过程也有大量的碳以 DIC、POC 和 DOC 的形式输入海洋,这部分受方法学制约鲜见报道。但已有个别研究结果显示:滨海湿地通过潮汐输入到海洋中的无机碳远超其沉积有机碳。因此,滨海湿地的实际年固碳能力,远超通过传统碳埋藏所估算的速率。
国际滨海湿地蓝碳研究热点
自 2011 年以来,国际上有关海岸带蓝碳生态系统固碳能力的研究已经逐渐成为热点;国内近几年亦有多个蓝碳研究项目在开展。Macreadie 等总结了蓝碳研究的 10 个重要问题,其中涉及滨海湿地蓝碳功能的问题的有 9 个,分别是:气候变化如何影响蓝碳系统的碳累积?人类干扰如何影响蓝碳系统的碳累积?蓝碳生态系统的分布及其时空格局如何?有机和无机碳循环过程如何影响碳排放?如何估算蓝碳系统中碳的来源?影响蓝碳系统中碳埋藏速率的因子有哪些?蓝碳系统和大气的温室气体交换速率如何?如何减少蓝碳估算中的不确定性?管理措施如何维持并提升蓝碳固碳功能?这 9 个问题既是当前滨海湿地蓝碳研究的热点,也是未来的主要研究方向。
总体上,我们对滨海湿地这一蓝碳生态系统的碳储量、速率、过程机制和生态系统服务功能尚缺乏足够的了解。以美国为例,尽管“蓝碳”这一概念是由美国科学家最先提出,但其研究人员对美国滨海湿地的蓝碳固碳速率的了解依然比较缺乏——已有的报道多集中在滨海湿地的碳储量,而对其碳通量和生态系统服务功能缺乏系统的总结 。为了弥补这一缺失,中国研究人员利用滨海湿地碳沉积数据和美国湿地调查数据,系统估算了美国当前国家尺度上的滨海湿地蓝碳系统碳埋藏能力;并利用联合国政府间气候变化专门委员会(IPCC)的气候模型预测数据和全球滨海湿地面积的模拟数据,建立了固碳速率与气候因子的经验模型,前瞻性提出:全球滨海湿地蓝碳系统的碳埋藏能力到 21 世纪末将持续增加。这是首次在国家(美国)和全球尺度上对滨海湿地蓝碳碳埋藏速率的系统估算与预测。遗憾的是,目前尚缺乏对中国滨海湿地固碳功能的系统模拟预测。
滨海湿地是全球变化的敏感区和脆弱区,其固碳功能如何响应人类活动和环境变化是当前蓝碳研究的热点。一方面,人类活动导致的生境破坏、环境污染极大地影响了滨海湿地的健康,从而影响其固碳功能。近几十年来,人口迅速增长和经济快速发展的需求使全球海岸带地区的土地利用发生着剧烈的变化。全球滨海湿地的碳汇功能和碳库储量在过去 1 个世纪显著降低。人为活动和环境变化,包括围垦、填海造陆、海堤建设等人为干扰,以及营养盐输入增加、气温升高等环境要素变化等,都将导致滨海湿地碳汇功能持续下降。以美国为例,由于人类活动的增加导致其滨海湿地面积比工业革命前下降超过50%。近些年,由于人类对滨海湿地生态系统碳汇功能的认识,如何保护滨海湿地资源,以及有效恢复受损湿地,已成为恢复生态学领域的重大科学问题。在滨海湿地恢复过程中,碳循环会发生重大变化,有效的生态恢复会降低湿地 CH4 排放,促进植物生长,从而提高有机质沉积速率,进而提高湿地的固碳和生态系统服务功能。除了人类活动导致的土地利用变化,在全球气候变化的背景下,滨海湿地受到富营养化、气温升高、海平面上升的多重压力,导致植物的生长与演替过程的变化,尤其是改变了与碳循环相关的生产与分解过程,最终影响其固碳功能 。来自美国盐沼湿地的长达 10 年氮肥增加模拟实验证实:海水中氮元素的增加会导致盐沼湿地退化 ,进而导致其碳汇功能降低。海平面上升是影响滨海湿地碳汇功能的另一个重要因素。通常,海平面上升会增加有机质的沉积;但是,当海平面上升速率超过滨海湿地沉积速率时,这些湿地会逐渐被海水淹没,导致其持续固碳功能的丧失。
中国滨海湿地的碳汇功能
中国拥有 1.8×104 km 的大陆海岸线,超过 2×106 km2的大陆架,分布着各种湿地类型。本文重点关注了盐沼、红树林和滨海滩涂湿地。
盐沼
盐沼湿地具备很强的固碳能力,其沉积物的碳埋藏速率平均约为 168 g C·m−2·a−1 。盐沼湿地是我国滨海湿地中面积最大的海岸带蓝碳生态系统类型,但因分类学差异导致其在总面积的统计多寡不一。Zhou 等估计我国盐沼湿地的面积为 1 207—3 434 km2;根据联合国环保署认可的全球盐沼湿地遥感数据,我国盐沼湿地的面积为 5 448 km2 ;叶思源 2017 年的调查数据显示我国有植被覆盖的滨海湿地和潮滩的总面积达 9 862 km2。然而,Mao 等[39]制作的我国最新的国家尺度湿地遥感图发现盐沼湿地面积仅为 2 979 km2。我国的盐沼湿地主要分布在环渤海湾、江苏省沿岸和长江口等地,在南方热带亚热带区域也有部分分布(表 1),其主要植被包括芦苇、盐地碱蓬等耐盐植物,以及互花米草等(图 1)。其中,互花米草是源自北美盐沼湿地的外来种,具备较强的适应性和耐受能力,是北美盐沼湿地的主要植被类型。我国在 20 世纪 80 年代广泛引种互花米草,用于滨海地区促淤造陆和保滩护岸等生态工程,但也导致其入侵光滩湿地、威胁本土植物和水鸟栖息地等生态问题。截至 2015 年,我国互花米草湿地面积达 546 km2,比 1990 年扩张了 502 km2,主要分布在江苏、浙江、上海和福建等地。这些新扩张的互花米草湿地主要侵占了原有的滨海滩涂,占比达 93%。
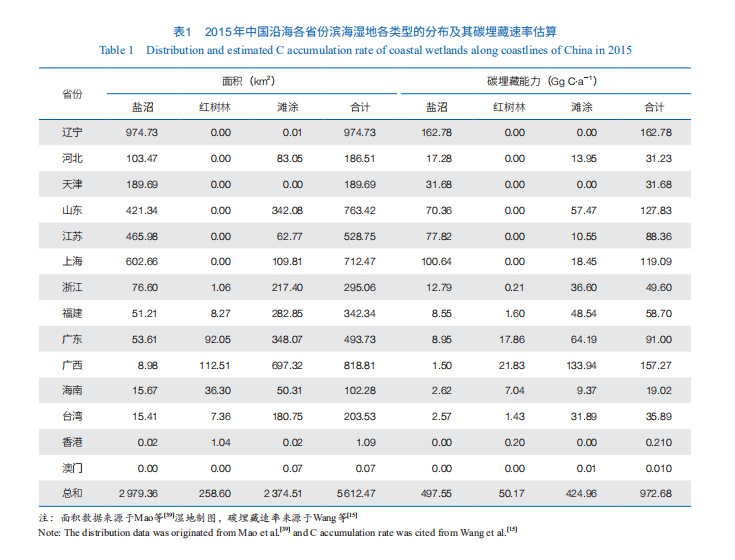
Wang 等利用盐沼湿地遥感数据并结合滨海湿地碳埋藏速率的实测数据,估算了我国盐沼湿地的碳埋藏速率为 1.19 Tg C·a−1。这一数据大于过去估算的数值0.26—0.75 Tg C·a−1,以及最近 Fu 等估算的 0.16 Tg C·a−1,主要原因是其采用的我国盐沼湿地面积(5 448 km2)较其他数据源大。本文采用较为保守的湿地面积(2 979 km2)计算,我国盐沼湿地年碳埋藏能力约为 0.50 Tg C·a−1(表1)。
红树林
红树林主要生长在热带和亚热带的海岸潮间带上,在全球尺度上红树林的总面积约为 1.4×105 km2,大于盐沼湿地的 6.23×104 km2。全球红树林每年沉积物碳埋藏速率约为 38.3 Tg C·a−1,这一数据远大于盐沼湿地的 12.6 Tg C·a−1。同时,红树林还可以向邻近海域输出 21 Tg C·a−1 的 POC 和 24 Tg C·a−1 的DOC[12]。因此,红树林被认为是固碳最有效的海岸带蓝碳生态系统。
我国红树林位于全球红树林分布的北缘,主要分布在广东、广西、海南和福建等省份。Wang 等估算了全球红树林的平均碳埋藏速率是 194 g C·m−2·a−1,而我国红树林的总碳埋藏速率约为 0.05 Tg C·a−1,这一数据与过去的其他研究相差不大,远小于我国盐沼湿地的碳埋藏速率,主要原因是我国现存红树林的面积过小。根据红树林面积遥感数据,2010 年我国红树林面积为仅为 171 km2 。但这一数值存在较大争议,Mao 等通过遥感制图得到的 2015 年全国红树林的总面积为 259 km2。根据国家林业和草原局的最新数据,我国红树林在过去 10 多年得到快速恢复,2020 年总面积已经有 289 km2,其中超过 70 km2 为近期新造和恢复的红树林。但即便按照最近的红树林面积数据,我国当前红树林面积也仅为历史上最高值(约为 2 500 km2)的 1/10 左右,恢复空间巨大。
滨海滩涂
滨海滩涂也是重要的海岸带生态系统类型,其主要包括泥质滩涂(Mudflat)、沙滩和基岩海岸等3 种类型,其中泥质滩涂具有相当强的碳埋藏能力。我国滨海滩涂的面积依据不同的遥感数据差异比较大。Mao 等制作的国家尺度湿地分布图显示我国 2015 年泥质滩涂的总面积为 2 374 km2,略低于我国的盐沼湿地总面积;而最近的卫星遥感工作显示我国滨海滩涂(含泥质滩涂、石滩、沙滩及部分浅海)的面积在 5 379—8 588 km2 之间。这些差异主要是由卫星影像的来源和拍摄频率所致。总体上,我国滨海滩涂的面积很大,甚至超过盐沼湿地和红树林的总面积之和,且以泥质滩涂为主,其沉积物埋藏速率高,具有很强的固碳潜力。这些滩涂沉积物埋藏碳的主要来源是周边的盐沼湿地和红树林的碳输入,以及海水中的 POC 和矿质结合碳组分的沉积,因此理应被纳入滨海湿地蓝碳碳汇计量体系。已有研究表明,泥质滩涂的沉积速率和碳埋藏能力与周边的盐沼湿地和红树林相当。我们根据最保守的滨海泥质滩涂的分布面积及其周围盐沼和红树林碳埋藏速率的数据,估算出我国滨海滩涂的碳埋藏速率下限约为 0.42 Tg C·a−1,这远高于我国红树林的碳埋藏能力,且仅次于盐沼湿地(表 1)。
此外,我国的滨海滩涂面临互花米草的入侵问题,在过去 30 多年间,近 467 km2 的滩涂演变为互花米草为主的盐沼湿地(图1)。相对于周边的盐沼湿地和红树林,滨海滩涂的生态系统净生产力比较低,仅为前者的 10%—20% 左右。互花米草入侵滩涂后不仅增加植物生物量和有机凋落物的输入量,而且其致密的植被可以减缓水流,加速沉积物的累积,提高沉积速率。此外,互花米草在滨海滩涂的定植吸收了大量的氮、磷等营养盐,能够减少陆地营养盐向近海富营养化海区的输入,提高近海初级生产力。总体上,互花米草入侵滨海滩涂其碳汇总量反而是增加的 ;若仅从碳汇视角看,这有利于我国滨海湿地生态系统固碳能力进一步提高 ;然而,其综合生态系统服务功能,仍需进一步评估。
发展方向及政策建议
滨海湿地具有很强的生态系统服务功能和碳汇价值,也具有很强自身恢复力。尽管面临人类活动干扰、海平面上升和气候变化等不利因素,全球滨海湿地的总面积在 21 世纪末仍会有一定程度的增加,其总的固碳能力,特别是碳埋藏速率会进一步增强。由于我国滨海湿地的沉积速率较高,如果没有人为对自然岸线的破坏和干扰,其在 21 世纪末的总面积仍会有较大比例增加,整体的碳汇和生态系统服务功能也会进一步增强。模型预测的结果显示,我国滨海盐沼和红树林湿地的碳埋藏能力在 21 世纪末可望增加到 1.82—3.64 Tg C·a−1。
但我国目前滨海湿地的总面积有限,过去几十年海岸带地区的滩涂围垦、鱼虾养殖、城市化及工业化等土地开发活动导致滨海湿地面积急剧减少,其固碳功能和碳汇潜力下降。1975—2017 年,我国天然湿地衰退率为 53.9%。因此,如何有效恢复滨海湿地,增加湿地面积,减少对湿地周围自然岸线的破坏,提高其自然恢复能力,增强现有滨海湿地的生态系统服务功能,对我国滨海湿地蓝碳功能的恢复和提高具有重要意义。
当前,我国急需加强滨海湿地的科学研究,保护其生态系统结构与服务功能的完整性,停止破坏性的滨海湿地开发活动,避免其蓝碳功能的快速损失,推进滨海湿地的生态恢复工作,重建和新建滨海湿地生态系统,恢复并增强其蓝碳功能,在保护自然的同时受惠于碳汇增益,让滨海湿地蓝碳为我国的碳中和战略作出更大贡献。因此,我们建议后续滨海湿地蓝碳科学研究和管理政策需着重加强下述 4 个方面。
建立海岸带生态系统野外观测研究网络。在全国范围内选择典型的海岸带生态系统,建立野外观测研究站,并纳入国家野外科学观测网络。通过多站点的联网观测,深入认识滨海湿地生态系统结构与服务功能,阐明其碳埋藏速率和温室气体排放的时空变化格局及其机制,并对滨海湿地中固碳能力较强的群落类型开展系统研究。
系统量化和预测我国滨海湿地蓝碳固碳功能。通过模拟人类活动和气候变化,结合地理信息系统和土地遥感数据,建立模型预测未来不同气候变化情景下蓝碳功能及其变化趋势,阐明我国滨海湿地对未来气候变化和人类活动的响应和适应机制,提高对我国滨海湿地蓝碳增汇机制的科学认识和对其未来碳汇强度的预测能力,突出其综合生态系统服务功能。
对外来种建立的滨海湿地开展系统研究。趋利避害,综合评估滨海湿地中外来种的生态风险和负排放效应;在合适的地区合理利用外来种恢复和新建盐沼湿地和红树林。
构建滨海湿地生态系统服务功能综合研究示范区。通过系统了解影响滨海湿地固碳功能的关键驱动因子,制定滨海湿地修复的法规和标准,研发相应的固碳增汇技术,在示范区建立适于不同滨海湿地的生态管理对策,实践我国滨海湿地生态系统服务功能最大化的生态管理方案。
(作者:王法明,中国科学院华南植物园小良热带海岸带生态系统研究站 南方海洋科学与工程广东省实验室 华东师范大学河口海岸学国家重点实验室;唐剑武,华东师范大学河口海岸学国家重点实验室;叶思源,中国地质调查局青岛海洋地质研究所;刘纪化,山东大学海洋研究院。《中国科学院院刊》供稿)